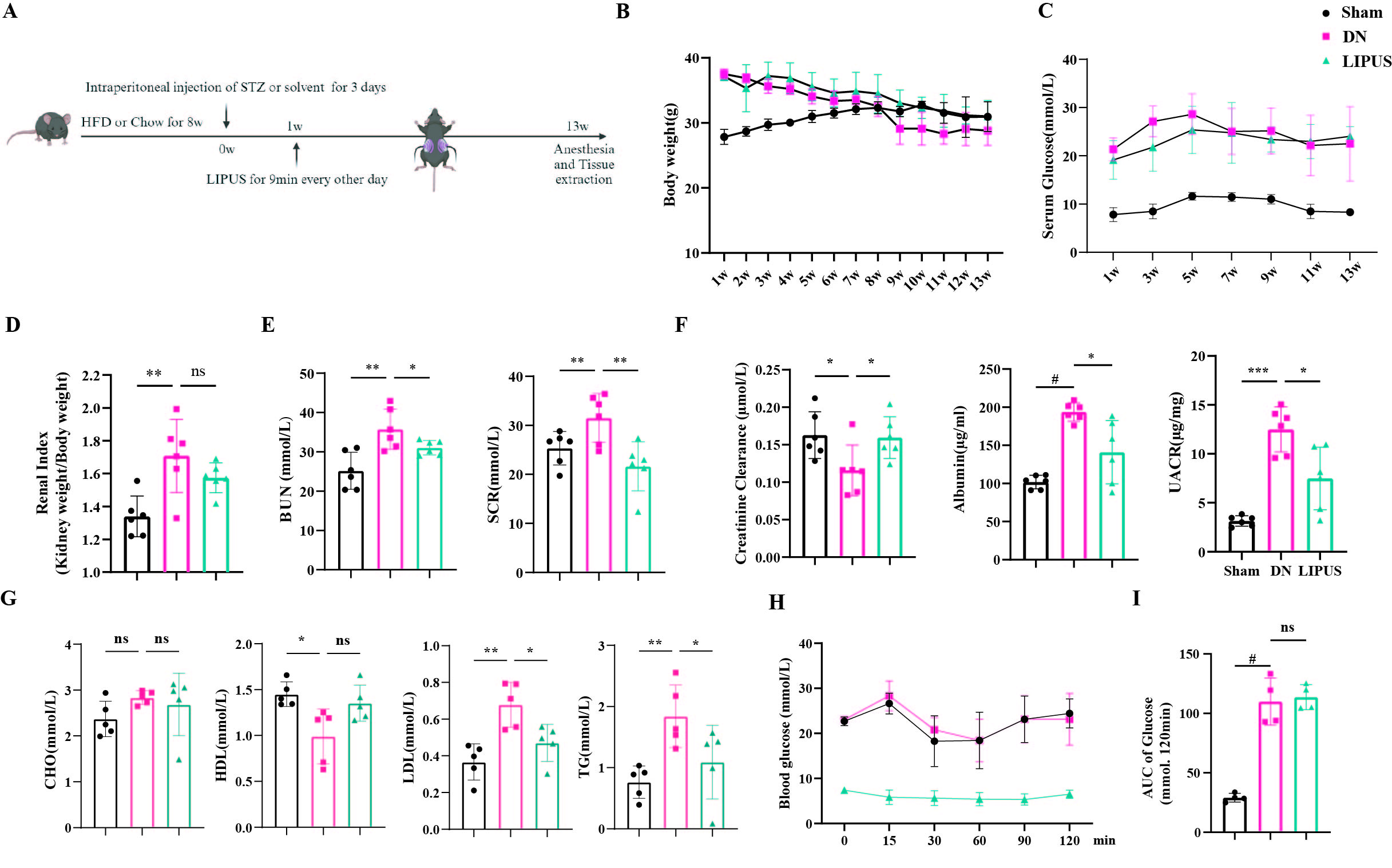
 As diabetes progressed, mice in both the DN and LIPUS groups
exhibited a continuous decrease in body weight, while blood glucose levels
remained elevated (Figure 1B and 1C). As shown in Figure 1D, DN mice displayed
renal hypertrophy, with a significantly higher renal index compared to the sham
group, and LIPUS treatment only partially ameliorate this condition. The
increased levels of blood urea nitrogen (BUN) and serum creatinine (SCR) in DN
mice were downregulated after LIPUS intervention (Figure 1E). Furthermore,
LIPUS treatment improved the lower creatinine clearance rate and attenuated the
increases in urinary albumin levels and the urinary albumin-to-creatinine ratio
in DN mice (Figure 1F). Notably, LIPUS also ameliorated lipid metabolism
disorders associated in DN mice, including low-density lipoprotein (LDL) and
triglyceride (TG) (Figure 1G). However, LIPUS treatment did not improve the
characteristic insulin resistance features of T2DM (Figures 1H and 1I). These
results suggest that the improvement of renal function by LIPUS in DN may be
independent of blood glucose management.
As diabetes progressed, mice in both the DN and LIPUS groups
exhibited a continuous decrease in body weight, while blood glucose levels
remained elevated (Figure 1B and 1C). As shown in Figure 1D, DN mice displayed
renal hypertrophy, with a significantly higher renal index compared to the sham
group, and LIPUS treatment only partially ameliorate this condition. The
increased levels of blood urea nitrogen (BUN) and serum creatinine (SCR) in DN
mice were downregulated after LIPUS intervention (Figure 1E). Furthermore,
LIPUS treatment improved the lower creatinine clearance rate and attenuated the
increases in urinary albumin levels and the urinary albumin-to-creatinine ratio
in DN mice (Figure 1F). Notably, LIPUS also ameliorated lipid metabolism
disorders associated in DN mice, including low-density lipoprotein (LDL) and
triglyceride (TG) (Figure 1G). However, LIPUS treatment did not improve the
characteristic insulin resistance features of T2DM (Figures 1H and 1I). These
results suggest that the improvement of renal function by LIPUS in DN may be
independent of blood glucose management.
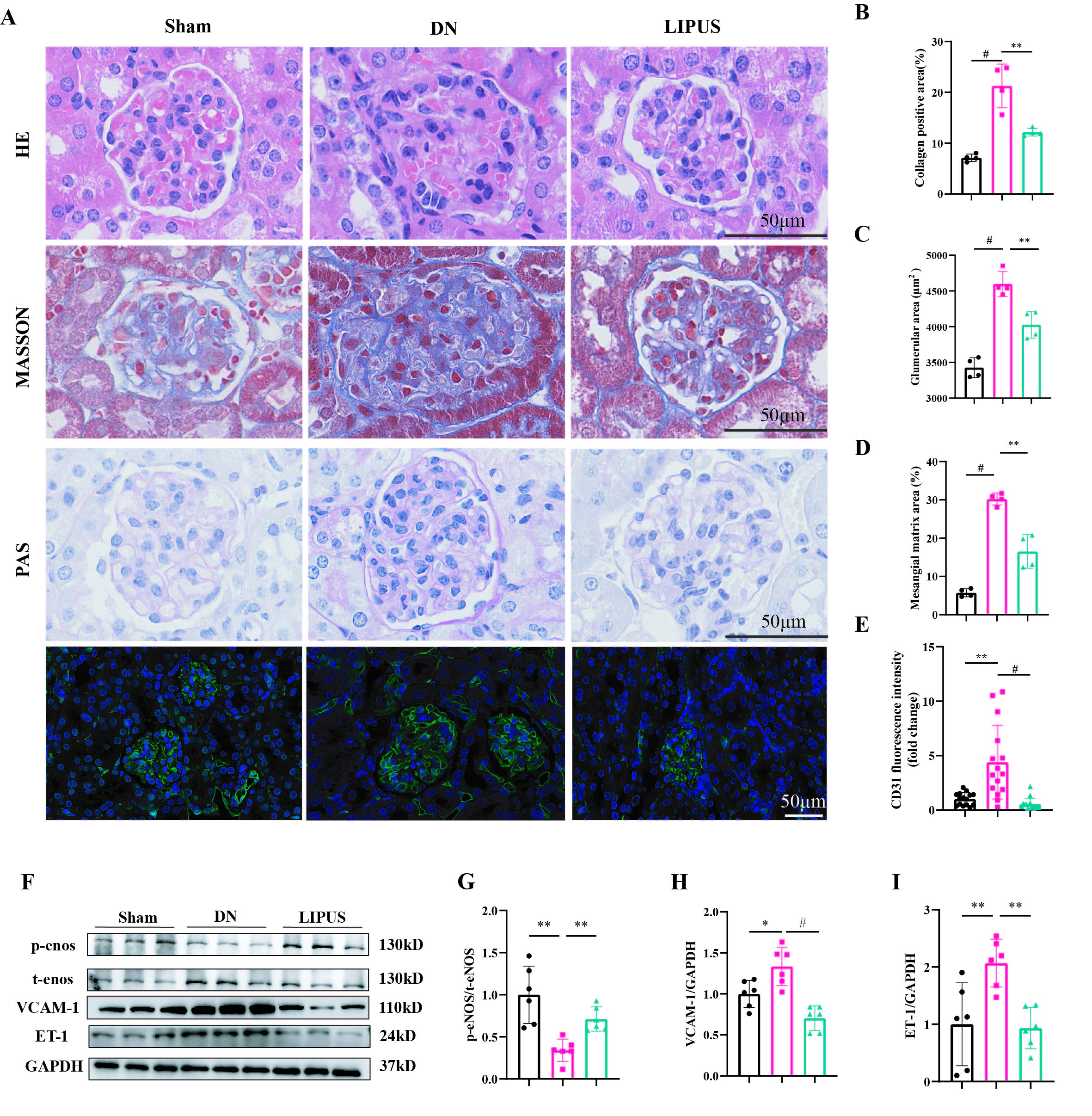
Furthermore, histological
examination was performed to evaluate the efficacy of LIPUS treatment. H&E staining
and Masson’s trichrome staining indicated that LIPUS effectively reduced the
extent of renal fibrosis compared with the DN group (Figures 2A and 2B).
Periodic Acid–Schiff staining revealed that DN mice exhibited significant
glomerular hypertrophy and mesangial matrix expansion, which were attenuated by
LIPUS treatment (Figures 2A-D). CD31 staining was used to assess GEC injury. In
the DN group, hypertrophic glomeruli were accompanied by abnormal vascular hyperplasia,
which was ameliorated following LIPUS intervention (Figures 2A and 2E). We
further examined markers associated with endothelial cell injury by Western
blot. Compared with the sham group, the DN group showed a significant decrease
in p-eNOS protein levels, along with increased expression of VCAM-1 and ET-1.
In contrast, LIPUS treatment upregulated p-eNOS expression and downregulated
the levels of VCAM-1 and ET-1 relative to the DN group (Figures 2F–2I). These
findings suggest that LIPUS treatment may confer a protective effect against
glomerular endothelial injury in DN.
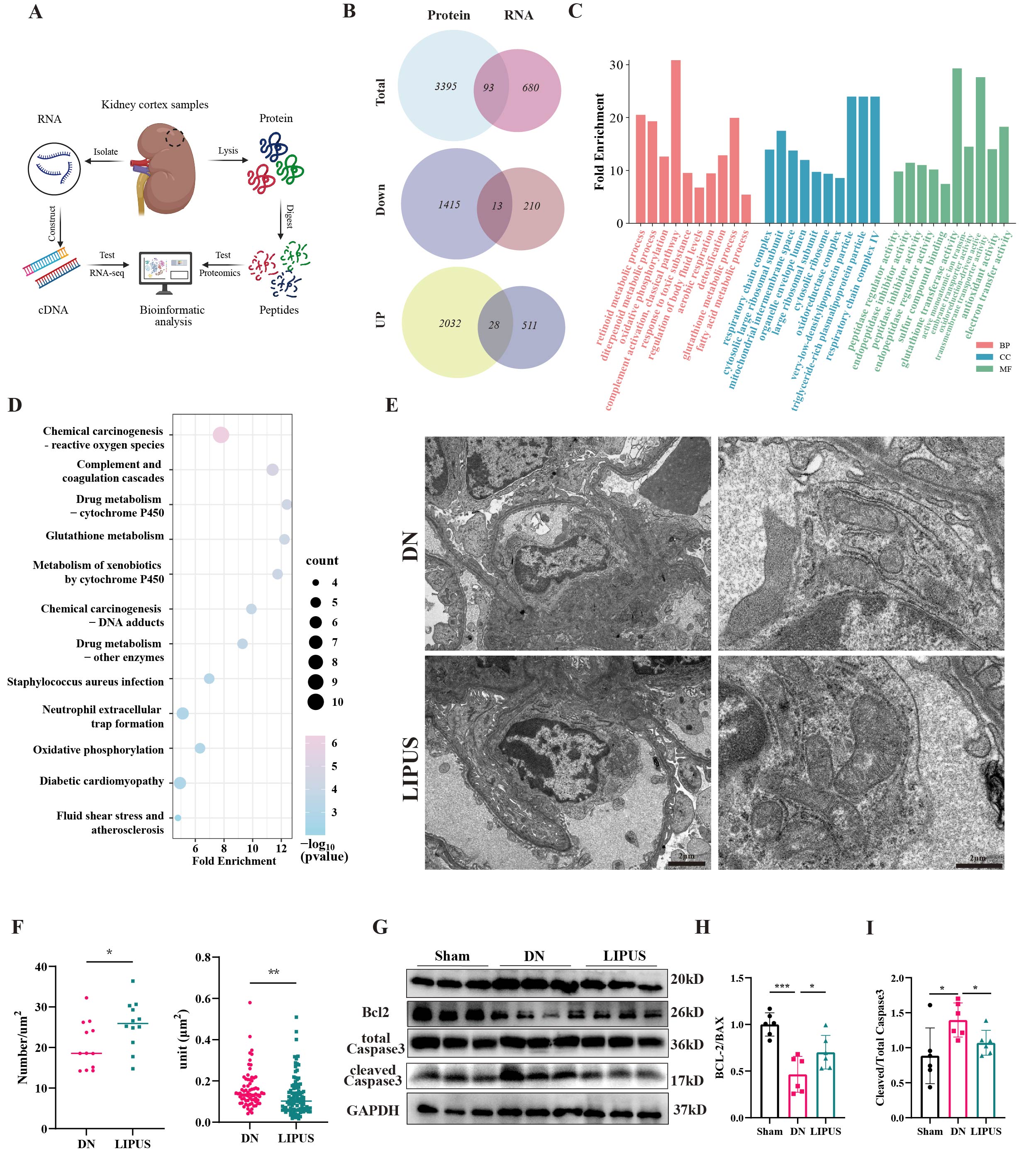
To further explore the underlying mechanisms and key targets of
LIPUS treatment, we performed 4D label-free proteomic analysis and RNA
sequencing on renal cortex samples from DN and LIPUS treated mice (Figure 3A).
Venn diagram analysis identified 93 common differentially
expressed genes (DEG) at the RNA and protein levels (Figure 3B). Among these
overlapping genes, the number upregulated by LIPUS was significantly greater
than those downregulated. Gene Ontology (GO) enrichment analysis revealed
multiple terms significantly associated with mitochondrial processes, including
oxidative phosphorylation, respiratory chain complex, mitochondrial
intermembrane space, and oxidoreductase complex (Figure 3C). Similarly, KEGG
pathway analysis indicated significant enrichment of mitochondria-related
pathways such as oxidative stress, cytochrome P450 metabolism, and oxidative
phosphorylation (Figure 3D). We next performed transmission electron microscopy
to evaluate mitochondrial ultrastructure in GECs. TEM images showed that LIPUS
treatment ameliorated the podocyte loss and glomerular structural damage
induced by DN (Figure 3E). Mitochondria in the GECs of DN mice exhibited a
significant reduction in number, accompanied by membrane disruption, structural
disorganization, and shortened, fragmented, and blurred cristae. In contrast,
LIPUS treatment effectively restored mitochondrial ultrastructure, increasing
mitochondrial number and reducing mitochondrial volume (Figure 3F). Consistent
with these morphological improvements, LIPUS upregulated the anti-apoptotic
protein BCL2 and downregulated the pro-apoptotic proteins BAX and cleaved
caspase-3, which are associated with the functions and integrity of mitochondrial.
These findings suggest that LIPUS may alleviate GEC dysfunction and injury by
attenuating mitochondrial damage.
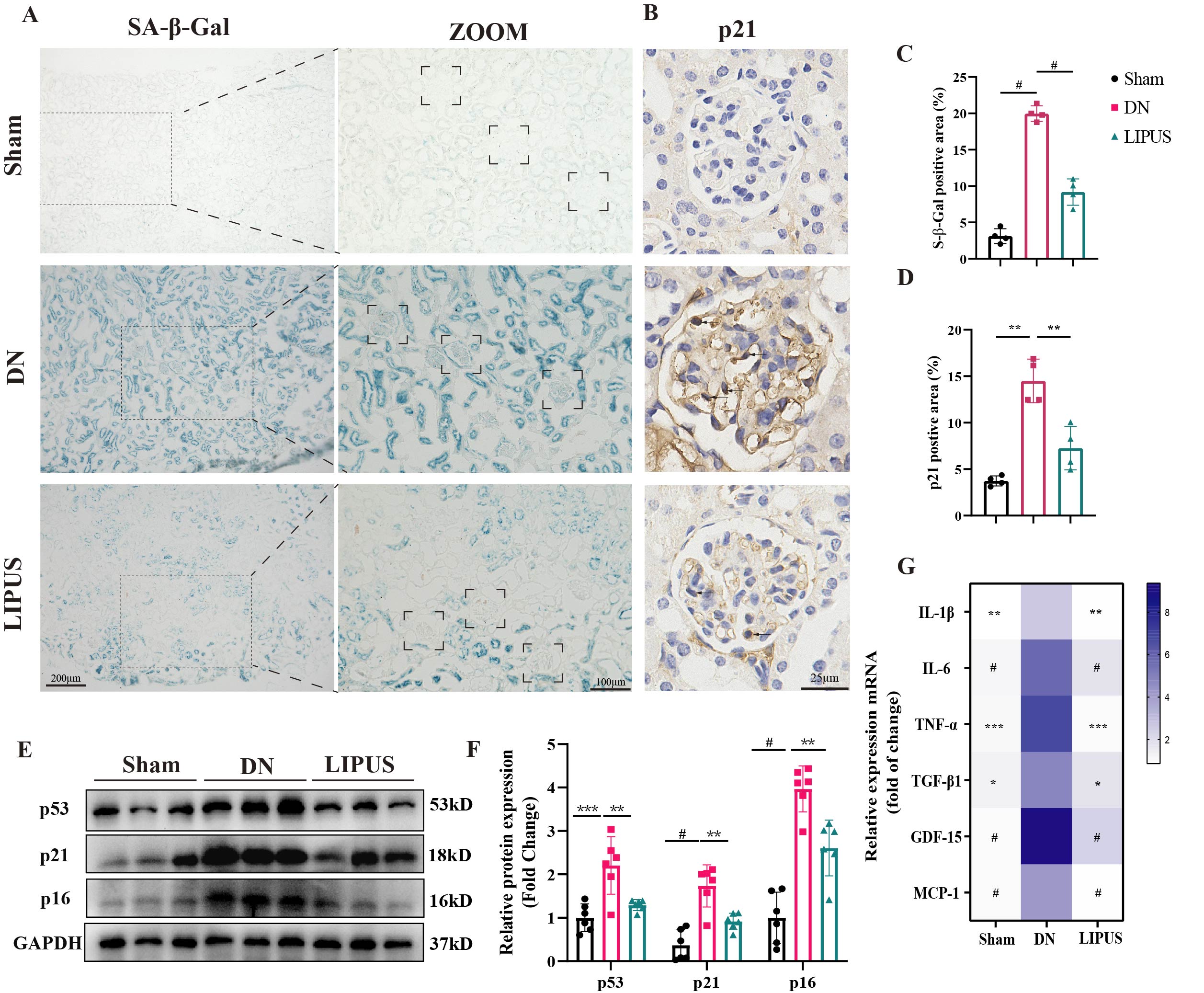
Given that
mitochondrial dysfunction is a pivotal factor of cellular senescence, and GEC
senescence has been established as a key pathological mechanism in diabetic
nephropathy, we next assessed whether the amelioration of mitochondrial injury
by LIPUS could subsequently attenuate the senescent phenotype in diabetic
kidneys. As shown in Figure 4A, renal sections from the DN group exhibited a
significantly larger SA-β-gal-positive area than those from the sham group, an
effect markedly attenuated by LIPUS treatment, particularly within glomeruli
(Figure 4C). Immunohistochemical staining for p21 in mouse glomeruli further
indicated that LIPUS reduced its elevated expression in DN kidneys (Figure 4B
and 4D). Consistent with these findings, Western blot analysis revealed that
the protein levels of senescence markers p53, p21, and p16 were upregulated in
DN kidneys compared with sham controls, and LIPUS treatment effectively
suppressed this increase (Figure 4E and 4F). Similarly, the mRNA levels of key
senescence-associated secretory phenotype (SASP) factors were significantly
elevated in DN kidneys relative to the sham group, and LIPUS treatment also
downregulated these SASP expression (Figure 4G).
